嵌合抗原受体T(chimericantigenreceptor-T,CAR-T)细胞是一种通过基因编辑表达跨膜嵌合抗原受体并具有特异性导向的T细胞。CAR-T作为一种新兴的免疫治疗,已在肿瘤治疗方面取得较大突破。近年来,基于当前自身免疫性疾病治疗的局限性,研究者们正在不断探索CAR-T及其衍生疗法在自身免疫性疾病中的潜在治疗价值。本文重点阐述CAR-T在自身免疫性疾病领域的研究进展及现阶段所面临的挑战,为自身免疫性疾病的精准治疗提供理论依据。
近年来,以嵌合抗原受体T(chimericantigenreceptor-T,CAR-T)细胞为代表的细胞免疫疗法吸引了越来越多的 自身免疫性疾病是一组免疫系统对自身抗原产生异常的免疫应答,导致自身组织细胞损伤或功能异常的临床病症。目前传统的治疗主要为非特异性的免疫抑制疗法,因其只能在体内建立短暂的免疫耐受,患者不仅需要长期维持治疗,且极大增加了感染及恶性肿瘤的风险。近年来,单克隆抗体疗法如抗CD20、抗肿瘤坏死因子-α(TNF-α)单抗等,逐渐被应用于多种疾病的临床治疗,它们能够靶向疾病特异性的抗原,减少机体全面免疫抑制的不良反应,然而仍无法长久性建立免疫平衡。CAR-T细胞的出现给自身免疫性疾病的治疗带来了新的希望。
01
CAR-T的结构
嵌合抗原受体(chimericantigenreceptor,CAR)是一种融合蛋白,由细胞内信号转导区(如CD3ζ)、跨膜区和胞外抗体单链可变区片段(singlechainfragmentvariable,scFV)组成[3]。CARs的特异性主要由胞外scFV区决定,它能够特异性识别肿瘤抗原,传递抗原信号,从而活化CAR-T细胞发挥其细胞毒效应(图1)。
第一代CARs的细胞内信号转导区只含有CD3ζ,不含共刺激结构域,由于反复暴露于抗原后无法有效诱导T细胞的扩增,其临床效益有限[4]。第二代和第三代CARs增加了共刺激结构域,细胞内信号转导区可与共刺激分子如CD28、CD和CD连接,促进了T细胞的活化和扩增,提高治疗效能[3]。最近,第四代CARs(又称TRUCKs)成功问世,它可以刺激非特异性免疫细胞(如NK细胞和巨噬细胞)应答,对因缺乏CARs识别目标而逃逸的肿瘤细胞发起攻击[5]。此外,第四代CARs还增加了安全开关和特定控制,如自杀基因[6-7],逻辑门控CARs和开关CARs等[8-10],实现了这一技术的高度可控。
图1左侧为CAR,右侧为嵌合自身抗体受体(chimericauto-antibodyreceptor,CAAR)的结构示意。两者的结构差异主要为胞外结构域,在CAR中,T细胞外的结构为抗体单链可变区片段,而CAAR为自身抗原片段。作用机制如下:CAR通过抗体单链可变区片段结合靶抗原,将信号传导入T细胞内,在细胞内信号转导区和共刺激域的作用下特异性地活化、扩增T细胞;CAAR则是通过自身抗原片段与致病性B细胞受体结合传导信号,从而抑制、消灭特异性B细胞,消除自身免疫病中的致病抗体产生。
02
自身免疫性疾病治疗范例及研究进展
2.1CAR-T细胞治疗自身免疫性疾病进展
2.1.1系统性红斑狼疮
系统性红斑狼疮(systemiclupuserythematosus,SLE)是一种由自身免疫介导的的慢性弥漫性结缔组织病。患者通常为生育年龄女性,以血清中出现多种自身抗体为特征,引起全身多脏器和组织受损。目前临床主要依靠非特异性免疫抑制治疗,如糖皮质激素及多种免疫抑制剂等,长期累积的副作用使得患者生活质量严重下降。近年来,利妥昔单抗(一种抗CD20的单克隆抗体)已成功治疗多种结缔组织病,如类风湿关节炎[11]、多发性硬化[12]等,但应用于系统性红斑狼疮的临床试验均未能获得满意的疗效,部分学者认为原因可能为无法完全耗竭产生抗体的B细胞或治疗时间过短[13],然而持续给药时患者的耐药性也随之增加,因此替代疗法亟需出现。
图1嵌合抗原受体和嵌合自身抗体受体的结构及作用机制
Kansal等[14]发现将表达CD19+嵌合抗原受体(CARs)的CD8+T细胞注入狼疮小鼠体内,小鼠血浆的CD19+B细胞被完全耗竭、致病抗体的滴度下降,有效改善了靶器官的功能(狼疮肾炎小鼠蛋白尿减轻或转阴、狼疮皮损及脱发缓解),并延长了狼疮小鼠寿命。此外,该实验中CAR-T细胞治疗的持久性也得到证实――单次给药即可产生长达一年的疗效。
最近,Jin等[15]将抗CD19CAR-T细胞分别于疾病发生前后注入SLE小鼠模型中,与对照组相比,抗CD19CAR-T细胞改善了SLE的器官受累症状,延长了小鼠的寿命,并能够有效预防SLE的发生。同时,他们还证实具有4-1BB共刺激域的CAR-T细胞比具有CD28共刺激域CAR-T细胞显示出更持久的B细胞耗竭。CAR-T细胞免疫治疗实现了小鼠体内产生致病抗体的B细胞的完全性、持久性耗竭,这为系统性红斑狼疮的治疗带来新的希望,目前进一步人体临床试验尚未开展。
2.1.21型糖尿病
1型糖尿病(type1diabetes,T1D)是一种在多种因素(自身免疫、遗传、病毒感染等)作用下胰岛B细胞受损,不能正常分泌胰岛素的自身免疫病。患者体内可有多种异常的自身抗体产生,如抗谷氨酸脱羧酶抗体、胰岛素抗体、胰岛细胞抗体等。目前临床主要依靠长期胰岛素维持治疗,由于患者体内胰岛素的绝对不足,易发生酮症酸中毒等严重并发症。
Zhang等[16]将针对1型糖尿病致病复合物I-Ag7-B:9-23(R3)的-CAR嵌入CD8+T细胞,再将-CAR-T细胞注入自发性非肥胖糖尿病小鼠模型体内,单次输注后即能显著降低小鼠血清胰岛素抗体滴度,延迟小鼠1型糖尿病的发生。在安全性方面,他们还评估了小鼠脾脏内CD4+T、CD8+T、B和NK细胞的比例,证明了CAR-T细胞治疗不会严重改变小鼠的免疫系统。然而,在持久性方面,他们发现小鼠体内的-CAR-T细胞随着时间延长逐渐减少,至25周时已无法检测,表明目前单次注入的CAR-T细胞治疗只能延迟,尚不能阻止1型糖尿病的发展。
2.2CAR-T细胞衍生疗法进展
2.2.1CAAR-T细胞免疫疗法
年,Ellebrecht等[17]提出了一种针对致病B细胞的特异性治疗模式――嵌合自身抗体受体T(CAAR-T)细胞免疫治疗,他们用致病性自身抗原片段代替CARs中的scFV,抗原片段能与B淋巴细胞上的特异性B细胞受体(B-cellreceptor,BCR)结合,通过杀伤特异性B细胞减少自身抗体的产生,而不引起广泛的免疫抑制(图1)。
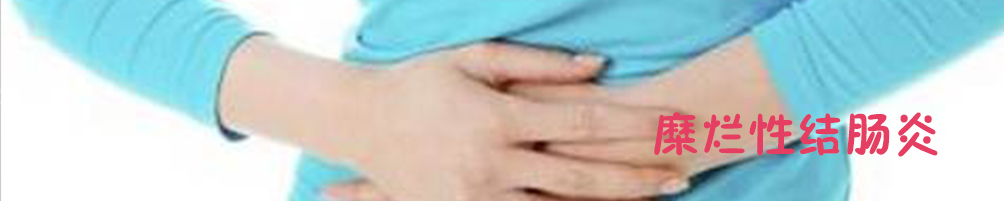
寻常型天疱疮(pemphigusvulgaris,PV)是一种严重的自身免疫性大疱性皮肤病,其发生是由于患者体内产生抗桥粒自身抗体(主要为抗Dsg-3抗体),干扰了角质形成细胞间的连接,导致棘层松解,从而形成水疱、大疱及粘膜损害[18](图2)。目前经典的治疗方法为糖皮质激素联合免疫抑制剂的治疗,在过去的几十年中虽使天疱疮的死亡率大幅下降[19],但也导致了严重副作用的发生[20]。
图2天疱疮患者躯干广泛糜烂、水疱
研究人员成功构建Dsg3-CAAR-T细胞,并将其注入PV小鼠模型体内。通过试验,他们发现CAAR-T细胞治疗后小鼠血清中的抗Dsg-3抗体滴度降低,且小鼠口腔粘膜水疱消失。同时,Ellebrecht等还证明CAAR-T细胞的存在不依赖于靶细胞,在靶细胞缺乏的情况下,3周后CAAR-T细胞可出现在脾内。另外,该实验在抗Dsg-3抗体的存在下重复进行,血清循环的自身抗体没有中和或消除CAAR-T细胞,这都证实了CAAR-T细胞在体内具有很强的持久性。然而,CAAR-T细胞在体内的确切存活时间尚未得到证实。在安全性方面,他们发现Dsg3-CAAR-T细胞只针对Dsg3特异性的BCR,而不清除全部B细胞,且Dsg3-CAAR-T细胞对角质形成细胞及其他正常组织均没有毒性,说明在小鼠模型中CAAR-T细胞不存在靶外毒性。
2.2.2CAR-Treg细胞免疫疗法
调节性T细胞(Regulatorycell,Treg)是一类控制机体自身免疫反应性的T细胞亚群,通过主动调节的方式抑制存在于正常机体内潜在的自身反应性T细胞的活化与增殖,参与维持机体免疫耐受。近年来,多克隆Tregs疗法应用于自身免疫性疾病的临床前及临床试验已初见成效,但也引起了非特异性免疫抑制的风险。基于此,研究人员将CAR嵌入Tregs细胞,形成抗原特异性Tregs,实现了靶向、精准治疗。
Fransson等[21]报道在多发性硬化的实验性自身免疫性脑脊髓炎模型小鼠中使用靶向特异性抗原的CARαMOG-Foxp3治疗,小鼠疾病症状减轻,且脑内IFN-γ和IL-12水平降低;Elinav等[22]将TNPscFv-CD28-FcRγ嵌入Treg治疗结肠炎,证明了CAR-Tregs在体内外均表现出明显的抑制效应,有效减轻了小鼠的结肠炎。除此以外,CAR-Tregs治疗包括自身免疫性肝炎、炎症性肠病、类风湿关节炎、自身免疫性糖尿病等在内的多种自身免疫性疾病的临床试验也正在进行中,未来可能成为自身免疫性疾病的潜在治疗方案。
03
挑战与应对
尽管CAR-T细胞治疗在小鼠体内取得了巨大的成功,但仍可能面临许多挑战,寻求这些问题的解决策略有助于推进其进一步的临床应用。
3.1治疗效能把控
3.1.1T细胞的效能
提高T细胞的效能对于治疗至关重要。首先,增加T细胞的剂量可以提高效能,但同时会相应增加毒性的风险[23-24],因此,治疗前需要确定合适的T细胞剂量,以维持两者之间的平衡,但这仍然是未知的。其次,T细胞初始亚群的选择也直接影响疗效[25]。据报道,CD4+和CD8+T细胞的结合可能带来协同效应[26]。而另一项研究发现在CAR-T的治疗中,仅CD4+T细胞的效果优于CD4+和CD8+T细胞的组合[27],关于T细胞亚群的选择还存在争议,需要进一步的研究。Ghassemi等[28]通过减少T细胞体外扩增时间限制其分化,证明了适当减少T细胞和CAR之间的间隔时间可以提高疗效。需要注意的是,在自身免疫性疾病中T细胞的功能常常受损,这可能影响治疗的效果。基于此,Meng等[29]选择NK细胞而不是T细胞作为嵌合细胞以提高疗效,但也带来了诸如移植物抗宿主病等问题。
3.1.2CAR及CAAR的设计
共刺激结构域在CAR-T细胞的功能中起着至关重要的作用,如CD28CAR-T和4-1BBCAR-T细胞均能够起到治疗作用,但在反应方式、不良反应、细胞因子和免疫抑制因子水平等方面存在差异[30]。研究表明,在CAR-T细胞中加入转基因特性可显著提高治疗的持久性[31-32],并且铰链可以增强特定CAR-T细胞的增殖和活性[33-34]。因此,为提升CAR-T细胞的活性,应充分考虑抗原靶点位置和铰链长度。
3.2不良反应
3.2.1脱靶效应
脱靶效应是指未能达到预先设定的目标,有所偏移而使得正常组织或器官受损的现象。在寻常型天疱疮的临床前试验中,尽管在小鼠模型上已经证明Dsg3-CAAR-T细胞在体内对携带桥粒的组织及正常组织没有毒性[17],但因存在个性化的因素(桥粒的膜间距离大于T细胞活化的最佳距离、嵌合的抗原表位截短等原因),尚不能说明CAAR-T及CAR-T细胞应用于其他自身免疫病治疗缺乏脱靶毒性。此外,Beatty等[35]利用mRNACAR-T细胞免疫治疗降低了脱靶毒性。
3.2.2细胞因子释放综合征
细胞因子释放综合征是CD19-CAR-T治疗过程中常见的毒性反应,主要是由于大量细胞因子如IL-6、TNF-α等释放所引起的全身性炎症反应[36-37]。在自身免疫性疾病治疗中,Ellebrecht等[17]认为CAAR-T细胞治疗寻常型天疱疮时不太可能出现这个问题,因为Dsg3-B细胞仅占体内B细胞的很少一部分;另一种观点是,当CAAR-T细胞被注射到PV患者体内,其皮肤受损、产生炎症反应时,炎症信号通路和先天性免疫因子可能导致严重的细胞因子释放综合征[38],进一步的研究还需开展。此外,一些生物标志物能够预测患者是否可能在细胞治疗过程中遭受严重的细胞因子释放综合征或神经毒性[39],因此它们可以成为这些毒性的早期信号。
3.2.3其他不良反应
除了以上常见的毒副作用,依然可能存在许多目前无法预知的不良反应在人体试验及临床应用中发生,因此设计“自杀”手段快速消灭CAR-T细胞的毒性至关重要。目前的方法主要利用生物基因工程技术,将生物蛋白融合到T细胞中,然后利用其对抗剂快速消灭[40-41]或使其自我凋亡[42]。
3.3治疗成本高昂
CAR-T细胞输注患者体内前需要进行活化、刺激及体外扩增,大多为个体化治疗方案,其所带来的时间与金钱成本高昂[43]。目前一些研究致力于通过基因工程的方法消除内源性T细胞受体[43-46],设计标准化商业产品,以解决细胞治疗中移植物抗宿主病的发生。Qasim等[47]报道了一种产生“通用”CAR-T的方法,即通过使用抗CD52抗体来沉默TCRA和CD52基因的表达,这些都为CAR-T日后的发展提供借鉴。
04
展望
近年来,细胞工程、基因编辑和细胞制造方面取得了巨大进展,使得细胞免疫治疗发展迅速,在许多疾病领域内发挥着越来越大的作用。CAR-T及其衍生细胞免疫疗法在临床前研究的基础上,已显示了初步满意的实验结果,有望进一步开展临床试验,为自身免疫性疾病的精准治疗提供新的希望。
预览时标签不可点收录于话题#个上一篇下一篇